| Neoaleurodiscus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Neoaleurodiscus | ||||||||||||
| Sheng H. Wu Sheng Hua Wu |
Neoaleurodiscus ist eine Pilzgattung aus der Familie der Schichtpilzverwandten (Stereaceae). Es handelt sich um eine Satelittengattung des Aleurodiscus-Komplexes, die erst 2010 durch Sheng H. Wu beschrieben wurde. Die Typusart Neoaleurodiscus fujii wurde an der Baumgrenze des Fuji (Vulkan) gesammelt, wo der Pilz am Stamm eines Rhododendrons wuchs. Die Gattung ist makroskopisch gekennzeichnet durch die scheibenförmigen Fruchtkörper und mikroskopisch durch perlschnurartig eingeschnürten Gloeozystiden, einfache, unverzweigte Hyphidien, knotig septierte Hyphen und die glatten, amyloiden und mehr oder weniger dickwandigen Basidiosporen. Acanthohyphidien kommen nicht vor.
Merkmale
BearbeitenMakroskopische Merkmale
BearbeitenDie Fuchtkörper sind scheibenformig bis effus-reflex. Die Hymeniumoberfläche ist glatt und der Rand der Rand ist scharf abgesetzt und oft etwas angehoben oder aufwärts gebogen.[1]
Mikroskopische Merkmale
BearbeitenDas Hyphensystem ist monomitisch. Die Hyphen selbst sind knotig sepiert, tragen also an den Septen Schnallen. Die zylindisch bis keulenförmigen Basidien sind viersporig. Neben den Basiden findet man im Hymenium als sterile Elemente moniliforme, also perlschnurartig eingeschnürte Gloeozystidien, die sich nicht mit Sulfobenzaldehyden anfärben lassen und einfache Hyphidien. Die ziemlich dickwandigen, amyloiden Basidiosporen sind fast kugelig bis breit ellipsoid und glatt.[1]
Systematik
Bearbeiten- Neoaleurodiscus-Stammbäume
-
 Abbildung 1: Maximum Likelihood-Baum der die gleichen Sequenzen verwendet wie Wu für die Berechnung Seines MP-Baumes. Die rot dargestellte Aleurodiscus aurantius-Sequenz fehlt in Wus MP-Baum. Alle weiteren Angaben zu dieser und den folgenden Abbildungen befinden sich in der jeweiligen Bildbeschreibung.
Abbildung 1: Maximum Likelihood-Baum der die gleichen Sequenzen verwendet wie Wu für die Berechnung Seines MP-Baumes. Die rot dargestellte Aleurodiscus aurantius-Sequenz fehlt in Wus MP-Baum. Alle weiteren Angaben zu dieser und den folgenden Abbildungen befinden sich in der jeweiligen Bildbeschreibung. -
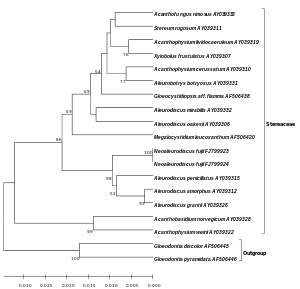 Abbildung 2 zeigt einen UPGMA-Baum, der mit Wus Sequenzen berechnet wurde. Hier bilden Neoaleurodiscus,
Abbildung 2 zeigt einen UPGMA-Baum, der mit Wus Sequenzen berechnet wurde. Hier bilden Neoaleurodiscus, -
 Abbildung 3: ZeitMaximum Likelihood-Baum
Abbildung 3: ZeitMaximum Likelihood-Baum



Wu berechnete aus 19 LSU-rDNA-Sequenzen von Vertretern aus der Familie der Schichtpilzverwandten (Stereaceae) einen Maximum Parsimony-Stammbaum (MP-Baum). Der Baum bestand auf der untersten Ebene aus zwei Hauptästen, zum ersten Ast gehören Arten, die nahe oder näher mit Stereum verwandt sind, zum zweiten Ast gehörten neben Neoaleurodiscus fujii, Aleurodiscus s.s. (Aleurodiscus grantii und Aleurodiscus amorphus), Aleurodiscus penicillatus, Acanthobasidium sowie Aleurodiscus weirii. Auf der zweiten Ebene spaltet sich der zweite Hauptast in zwei Unteräste auf. Den ersten Unterast bildete Neoaleurodiscus und Aleurodiscus aurantius (A. aurantius-Sequenz fehlt in Wus MP-Baum), zum zweiten Unterast gehörten die übrigen Arten. Diese spalten sich auf der 3. Ebene wiederum in zwei Unteräste auf. Der erste Unterast wird von Aleurodiscus s.s. und er zweite von Aleurodiscus penicillatus, Acanthobasidium sowie Aleurodiscus weirii gebildet.
Abbildung 1 zeigt einen Maximum Likelihood-Baum (ML-Baum), für dessen Berechnung die gleichen Sequenzen verwendet wurden, die auch Wu für seinem MP-Baum verwendet hat. Zusätzlich wurde eine Sequenz für Aleurodiscus aurantius (in der Abbildung rot dargestellt) hinzugefügt, . Er stimmt in den meisten Punkten mit Wus MP-Baum überein, nur das der 2. Unterast auf der 3. Ebene wiederum mit Aleurodiscus s.s., A. penicillatus und Acanthobasidium/A. weirii drei Unteräste hat. Berechnet man aber mit den gleichen Sequenzen einen UPGMA-Baum so wandert der Acanthobasidium an die Basis der Stereaceae und Neoaleurodiscus/A. aurantius, Aleurodiscus s.s. und A. penicillatus bilden ein Monophyllus. Verwendet man mit Peniophora cinerea und Hyphodontia alutaria eine entferntere Outgroup und fügt ein paar weitere Sequenzen hinzu (in der Abbildung 3 rot dargestellt), so erhält man einen ML-Baum, der den UPGMA-Baum (Abbildung 2) zu bestätigen scheint.
Allerdings bleiben die genauen Verwandschaftsverhältnisse zwischen den genannten Arten unklar. Zum einen, weil man zu wenig DNA-Sequenzen hat um einen verlässlichen Baum zu berechnen, zum anderen, weil LSU-Sequenzen zwar gut geeignet sind um Verwandtschaftsverhältnisse auf Familienebene aufzudecken, aber kaum geeignet sind, um die Verwandtschaftsverhältnisse von nahe verwandte Arten aufzulösen.
| Merkmal | Aleurodiscus amorphus[2] |
Aleurodiscus grantii[3] |
Neoaleurodiscus fujii[1] |
Aleurodiscus aurantius[4] |
Aleurodiscus penicillatus[5] |
Aleurodiscus monilifer[6][7] |
|---|---|---|---|---|---|---|
| Fruchtkörper -form |
Scheiben- bis schüsselförmig | Scheiben- bis schüsselförmig | Scheibenförmig oder effuso-reflex | Effus oder corticioid | Effus oder corticioid | Scheibenförmig |
| Fruchtkörper- dicke in mm |
0,5–1 | 0,3–1 | 0,25–0,6 | 0,1–0,4 | 0,1–0,4 | 0,2–0,5 |
| Schnallen | nein | ja | ja | nein | Ja/nein | ja |
| Gloeozystidenform | moniliform | moniliform | moniliform | moniliform | moniliform | moniliform |
| Gloeozystiden in µm |
110–220 x5–10 | – × 4–7 | 80–150 × 5–10 | 40–60 × 12–20 | 35–65 × 5–12 | 60–120 × 7–11 |
| Hyphidien | Unverzweigt bis wenig verzweigt | Unverzweigt bis wenig verzweigt | Unverzweigt | Dendrohyphidien | Unverzweigt, , Acanthohyphidien | Unverzweigt bis wenig verzweigt |
| Basidien | Zylindrisch bis keulig | Zylindrisch bis keulig | Zylindrisch bis keulig | Zylindrisch bis keulig, | keulig | keulig |
| Basidienmaße in µm |
80–200 × 18–25 | 150–250 × 25–28 | 110–130 × 4–18 | (30) 50–80 (95) × 10–15 | 55–90 (110) × 9–12 | (60) 70–80 × 10-12 |
| Basidiosporen | fast kugelig - ellipsoid | fast kugelig - ellipsoid | breit ellipsoid bis fast kugelig | ellipsoid bis fast kugelig | fast kugelig, ellipsoid | ellipsoid |
| Ornamentierung | stachelig | stachelig | glatt | Fein stachelig | Fein stachelig | glatt |
| Sporenmaße in µm |
(20) 24–28 (35) × (15) 18– 22 (25) | 22–32 (39) × 18–24 (28) | 19–24 × 14,5–18 | (14) 16–20 (22) × (10) 12–15 (16) | 15–20 (25) × 13–19 (21) | 22–26 × 14–17 |
| Substrat | Nadelbäume | Nadelbäume | Angiosperme | Angiosperme (selten auch auf Eibe) | Nadelbäume | Nadelbäume (Zedern) |
Vorerst bleibt unklar, ob Neoaleurodiscus/A. Aurantiacus nur einen Ast innerhalb von Aleuodiscus s.s. bilden oder ob sie, wie es Wu verschlägt ein eigenständiges Taxon bilden. Das gleiche gilt für A.
Eine Analyse der LSU nrDNA und auch der 16 zeigt, dass Aleurodiscus aurantius sehr nahe verwandt ist und mit Neoaleurodiscus fujii eine Abstammungslinie (Klade/Monophyllum) bildet. Eine Tatsache auf die Wu in seiner Beschreibung nicht hin weist. Neben der Ähnlichkeit der LSU nrDNA gibt es auch morphologisch einige Ähnlichkeiten zweischen beiden Arten. Die großen fast kugeligen Sporen, die moniliformen, sulfoaldehydneagiven Gloeozystiden ... Aufgrund von molekularbiologischen Daten ist Aleurodiscus amorphus, Aleurodiscus grantii ebenfalls nahe verwandt. Laut Wu unterscheidet sich Neoaleurodiscus von den beiden anderen Gattungen durch die glattwandigen Basidiosporen und von Aleurodiscus (s.s.) durch seine knotig-septierten Hyphen (Hyphen mit Schnallen). Allerdings gibt es auch bei Aleurodiscus grantii Hyphen mit Schnallen. Von Acanthobasidium unterscheidet er sich durch das Fehlen von Acanthohyphiden.
Arten
BearbeitenDie Gattung beherbergt zwei Arten. Neben der Typusart stellte der Autor Aleurodiscus monilifer Malençon, ohne dies durch molekularbiologische Daten zu stützen.
Ökologie und Verbreitung
BearbeitenDie Typusart Neoaleurodiscus fujii wurde an der Baumgrenze des Fuji (Vulkan) gesammelt, wo der Pilz am Stamm eines Rhododendrons wuchs. Aleurodiscus monilifer wächst im Atlasgebirge (Marokko) auf der Rinde der Atlas-Zeder (Cedrus atlantica). [1][7]
Quellen
Bearbeiten- Neoaleurodiscus. Sheng H. Wu, Mycologia 102 (1): 218 (2010). In: MycoBank.org. International Mycological Association, abgerufen am 30. Juli 2014.
- J. Boidin, J. Mugnier & R. Canales: Taxonomie moléculaire des Aphyllophorales. In: Mycotaxon. Band 66, 1998, S. 464 (cybertruffle.org).
Einzelnachweise
Bearbeiten- ↑ a b c d Sheng-Hua, Dong-Mei Wang, and Shi-Yi Yu: Neoaleurodiscus fujii, a new genus and new species found at the timberline in Japan. In: Mycol. Soc. America (Hrsg.): Mycologia. Band 102, Nr. 1, 2010, S. 217–223 (mycologia.org [PDF]).
- ↑ Aleurodiscus amorphus in der CBS Aphyllophorales Datenbank. In: cbs.knaw.nl. 1997, abgerufen am 24. März 2013.
- ↑ Aleurodiscus grantii in der CBS Aphyllophorales Datenbank. In: cbs.knaw.nl. 1997, abgerufen am 24. August 2014.
- ↑ Aleurodiscus aurantius in der CBS Aphyllophorales Datenbank. In: cbs.knaw.nl. 1997, abgerufen am 24. März 2013.
- ↑ Aleurodiscus penicillatus in der CBS Aphyllophorales Datenbank. In: cbs.knaw.nl. 1997, abgerufen am 24. August 2014.
- ↑ Aleurodiscus monilifer in der CBS Aphyllophorales Datenbank. In: cbs.knaw.nl. 1997, abgerufen am 24. August 2014.
- ↑ a b Maria Nunez und Leif Ryvarden: Aleurodiscus monilifer. In: The genus Aleurodiscus (Basidiomycotina). In: Synopsis fungorum. Vol: 12. Fungiflora, Oslo, Norwegen 1997, S. 107 (mycobank.org).
Weblinks
Bearbeiten
Die Neoaleurodiscus fujii Sheng H. Wu 2010, Ascomycota Neoaleurodiscus monilifer (Malençon) Sheng H. Wu 2010, Ascomycota