Schwarzer Seeigel
Der Schwarze Seeigel (Arbacia lixula) ist ein kleiner benthisch lebender Seeigel der Küstenregion (Litoral). Er ist der Gattung Arbacia innerhalb der Familie der Arbaciidae zugehörig.
| Schwarzer Seeigel | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Arbacia lixula am Mittelmeer | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Arbacia lixula | ||||||||||||
| (Linnaeus, 1758) |
Namensgebung
BearbeitenLinné beschrieb den Schwarzen Seeigel 1758 erstmals in seiner 10. Ausgabe der Systema Naturae als Echinus lixula. Die Zuordnung zur Gattung Arbacia erfolgte 1835 durch John Edward Gray. Der Gattungsname soll sich vom Namen Arbaces, einem Nebencharakter des historischen Gedichtes Sardanapalus von Lord Byron, das 1821 veröffentlicht wurde, ableiten. Der Artname stammt nach Harvey (1956) womöglich vom Namen eines flachen, runden Gebäcks, das im alten Rom aus Mehl und Käse hergestellt wurde und selbst seine Namensgebung vom lateinischen Verb lixare (lat.: „in Wasser kochen“) erhielt[1].
Merkmale
BearbeitenAls einer der wenigen heute vorkommenden (rezenten) Vertreter der Ordnung der Arbacioida zeichnet sich der Schwarze Seeigel zunächst durch Merkmale aus, die allen „regulären“ Seeigeln (Regularia) gemein sind. Sein radiärsymmetrischer Körper weist eine streng pentamere Form auf. Mund und After des Schwarzen Seeigels befinden sich an den beiden Körperpolen des runden, jedoch seitlich leicht abgeflachten Körpers[2][3]. Weiterhin besteht das halbkugelige (hemisphärische) innere Kalkskelett (Testa) aus 5 Doppelreihen von Ambulakralplatten. Zwischen diesen liegen 5 Doppelreihen von Interambulakralplatten, die die Stachelbasen aufweisen. Die paarigen Ambulakralreihen sind den Armen der Seesterne homolog und weisen Poren auf, durch die die Ambulakralfüßchen nach außen münden. Diese Platten sind starr, fest miteinander verbunden und enden aboral am Analfeld (Periprokt) an dem der After ausmündet. Die fünf Terminalplatten (Endplatten) dieser Doppelreihen bilden einen festen Ring um das Analfeld. Am Terminalporus der Testa treten die Radiärkanäle des Hydrocoels durch diese Terminalplatten aus. Die Kalkplatten der Testa bilden auf der Oralseite einen Ring um das Mundfeld (Peristom), aus dessen Mitte die Zähne der Laterne des Aristoteles, dem Kauapparat der Seeigel, ragen. Diese besteht aus 5 pfeilförmigen, radiär angeordneten Pyramiden („Kiefern“), die jeweils einen permanent nachwachsenden Zahn tragen. Dieser Komplex ist im Peristom über Muskeln und Bänder aufgehängt, die vom Innenrand des Peristoms ausgehen und als Heber und Senker agieren. Sie führen nicht nur die Zähne selbst, sondern können die gesamte Laterne aus dem Mund stülpen und einziehen oder sie innerhalb der Mundöffnung hin und her bewegen. Im entspannten Zustand ragen die Zähne leicht aus der Mundöffnung heraus[2]. Das Peristom trägt eine weiche Bindegewebsschicht aus welcher die Mundfüßchen des Schwarzen Seeigels austreten[2]. Hierbei handelt es sich um Ambulakralfüßchen, die an ihren Enden mit kleinen Saugnäpfen bestückt sind. Diese dienen dem Schwarzen Seeigel zur Anheftung an vorzugsweise hartem und glattem Substrat. Die Primärstacheln dieser Art sind gleichmäßig dicht in Reihen entlang der Ambulacren und Interambulacren angeordnet. Sie sind brüchig und sehr spitz zulaufend, etwa 3 cm lang und mit einem Bindegewebsband, durch welches sie bewegt werden, auf den nicht perforierten Tuberkeln (Gelenkhöcker) der Testa verankert. Sie können bei Berührung sofort aufgerichtet werden[3] und dienen dem Schutz vor Prädatoren sowie der Fortbewegung. Diese Art besitzt allerdings keine Sekundärstacheln[4]. Da die Ambulakralfüßchen auf der Aboralseite des Schwarzen Seeigels im Vergleich zu anderen Vertretern der Regularia nicht mit Saugnäpfchen bestückt sind[3], ist dieser in seiner Fortbewegung sehr eingeschränkt und besitzt nicht die Fähigkeit sich zu maskieren, d. h. lebende Tiere tragen keine Fremdkörper auf ihrer Aborseite[5][2]. Die typischen gestielten, trifoliaten Greifarme (Pedicellarien) sind allen Echinodermata gemein und dienen A. lixula zur Sauberhaltung des Körpers d. h. beispielsweise zur Entfernung kleiner Partikel oder Larven anderer Meerestiere, die sich zwischen den Stacheln einnisten[2][6]. Insgesamt weist die bilateralsymmetrische, oral abgeflachte Testa einen maximalen Durchmesser von 6 cm auf und ist ausgenommen der Stacheln von einem schwarzen Epithel überzogen. Die nackte Testa ist rötlich bis violettbraun, wodurch die Ambulacren durch ihre Lage zwischen zwei dunkleren Streifen, die außerdem die Lage der Poren anzeige hervorgehoben werden[3]. Das lebende Individuum zeichnet sich durch seine dunkelbraune bis tiefschwarze Färbung aus. Oftmals lässt sich beobachten, dass die Stacheln der Oralseite eher hellbraun gefärbt sind. Ebenso zeichnen sich kleinere Individuen meist durch Färbung aus[7]. Der Bau der Gonaden des Schwarzen Seeigels ist sehr typisch für alle Eleutherozoen. Sein Hämal- und Nervensystem sind dem der Seesterne ähnlich[6].
-
 Aboralseite der nackten Testa von A. lixula
Aboralseite der nackten Testa von A. lixula -
 Außenansicht der Laterne des Aristoteles
Außenansicht der Laterne des Aristoteles -
 A. lixula im Profil
A. lixula im Profil -
 Peristom von A. lixula
Peristom von A. lixula




Ökologie
BearbeitenLebensraum und Verbreitung
BearbeitenDas geographische Vorkommen von A. lixula beschränkt sich auf Spanien und Portugal, das Mittelmeer, das östliche Südamerika, die Macaronesischen Inseln, die Atlantikküste und Westafrika[8], wobei er am Mittelmeer stark zunehmende Bestände aufweist[3]. Aufgrund von Fossilienfunden sagt man A. lixula einen neotropischen Ursprung nach, da die Schließung des Isthmus die pazifische Art A. stellata von der atlantischen Verwandtschaftsgruppe (A. punctulata / A. lixula) getrennt zu haben scheint. Demnach sollen A. lixula und die karibische Art A. punctulata Schwesterarten sein, die sich nach der Schließung dieser Landenge schnell voneinander wegentwickelten[1]. Untersuchungen lassen außerdem darauf schließen, dass der Schwarze Seeigel das Mittelmeer trotz seiner Häufigkeit erst innerhalb des letzten Jahrhunderts besiedelte[1]. Er ist als Vertreter der Regularia ein typischer Bewohner des Benthos und einer der häufigsten Vertreter der Seeigel auf Küstenfelsen des Sublitorals, wo er sich bevorzugt auf mit Algen bedeckten Kalksteinen aufhält. Man findet ihn typischerweise vom Flachwasser bis hin zu einer Tiefe von ungefähr 30 m, wobei er vereinzelt auch noch in Tiefen bis zu 50 m vorkommt[7]. Neben dem Steinseeigel (P. lividus) kolonisiert er Kahlgebiete und Ansammlungen von Makroalgen, wo er sich bevorzugt auf krustenbildenden Kalkalgen niederlässt und durch seine Ernährungsweise zum Erhalt dieser Kahlgebiete beiträgt[9][10]. In Algenwäldern findet man ihn sehr selten. Bisher ist nicht eindeutig ob der Schwarze Seeigel diese meidet, da er krustenbildende Algen als Nahrung bevorzugt, oder ob die Wahl seines Lebensraumes auf seiner Unfähigkeit zur Fortbewegung in Algenwäldern beruht[10]. Diese Art ist zwar nicht gesteinsbohrend, kann sich aber durch die Ambulakralfüßchen auf seiner Oralseite in Brandungszonen stark ans Gestein ansaugen[11]. Man findet ihn allerdings nicht nur auf vertikalen, felsigen Oberflächen, auf denen er einer starken Hydrodynamik ausgesetzt ist, sondern auch auf flachen, felsigen Gründen, zerbrochenen Felsblöcken und selten sogar Sand[1].
Ernährung
BearbeitenTraditionell wurde der Schwarze Seeigel aufgrund der Analyse seines Mageninhaltes als Pflanzenfresser (Herbivor) angesehen. Weitere Untersuchungen zeigen jedoch, dass es sich bei dieser Art um einen Allesfresser (Omnivoren) mit Tendenz zur Karnivorie handelt[10]. Aufgrund seiner eingeschränkten Mobilität ernährt sich A. lixula von sessilen oder sehr langsamen, nicht jedoch von freischwimmenden (pelagischen) Organismen[4]. Beim Abweiden seiner Nahrung macht er sich die Zähne seines Kauapparates zum Lösen und Abschaben seiner Nahrung vom Substrat zu Nutzen. Er ernährt sich unter anderem von verschiedenen Gattungen der Braunalgen, wie Cystoseira spp. oder den Vertretern der Dictyotales[12][13]. Bevorzugt frisst er allerdings selbst in Algenwäldern krustenbildende Kalkalgen[8], weshalb ihm oftmals ein selektives Fressverhalten nachgesagt wird[10]. Darüber hinaus ernährt sich A. lixula beim „Weidegang“ auch von sessilen Invertebraten, aufgrund dessen diese Art nach Wangensteen auch als „Wolf im Schafspelz“ bezeichnet wird[1]. Es lässt sich daher auch vermuten, dass seine Vorliebe für krustenbildende Kalkalgen auf den mit diesen assoziierten Invertebraten beruht.
Lebensgemeinschaften
BearbeitenViele Vertreter der im Litoral oder benthisch lebenden Arten leben eng vergesellschaftet mit A. lixula und suchen Schutz in dessen direkter Nähe. So halten sich Jungfische der Gobiidae (Grundeln), Gobiesocidae (Ansauger), Blennidae (Schleimfische) und Tripterygiidae (Dreiflossen-Schleimfische) unter oder in unmittelbarer Nähe dieses Seeigels auf. Crustaceae (Krebstiere) wie Garnelen, Amphipoden (Flohkrebse) und Paguroidea (Einsiedlerkrebse) oder auch Polychaeten wie Schuppenwürmer profitieren vom Schutz zwischen seinen Stacheln. Weichtiere wie Käferschnecken und Vorderkiemerschnecken findet man meist unter dem Schwarzen Seeigel[14].
Prädatoren
BearbeitenFische zählen zu den wichtigsten Prädatoren der Seeigel. So gelten Diplodus sargus (Geißbrasse) und Diplodus vulgaris (Zweibindenbrasse) zu den wichtigsten Arten unter den Prädatoren des Schwarzen Seeigels während Coris julis (Meerjunger) als Vertreter der Labridae (Lippfische) nur juvenile Individuen erbeutet. Je nach Größenverhältnis zwischen Räuber und Beute machen sich die beiden Diplodus-Arten unterschiedliche Fressgewohnheiten zu Nutze. Kleinere Individuen (<1 cm) können durch Ansaugen im Ganzen verschlungen werden, während größere Individuen an ihrer Basis vom felsigen Substrat gelöst und umgedreht werden, sodass ihre Testa durch Bisse ins Mundfeld aufgebrochen werden kann. Den Arten Chromis chromis (Mönchsfisch), Thalassoma pavo (Meerpfau), Serranus cabrilla (Sägebarsch) und Parablennius gattorugine (Gestreifter Schleimfisch) kann zwar nicht das Räubertum an sich nachgewiesen werden, doch fressen sie tote Exemplare dieser Art. Vor allem Chromis chromis kann beim Fressen der im Wasser treibenden Fragmente von A. lixula beobachtet werden. Da die beiden Vertreter der Gattung Diplodus als zugehörige der Familie der Sparidae (Meerbrassen) einen wichtigen ökonomischen Nutzen tragen, nimmt die Überfischung jener Arten einen Einfluss auf die Bestände von A. lixula. Die Auswirkungen dieser Prädatoren und Aasfresser reicht über sogenannte trophische Kaskaden weit über ihren direkten Einfluss auf die Populationen von A. lixula hinaus und kann eine nachhaltige Veränderung des Benthos bewirken[15][16].
Fortpflanzung und Larvalentwicklung
BearbeitenBei A. lixula handelt es sich um eine getrenntgeschlechtliche Art, deren Sexualdimorphismus auf die Längenunterschiede der Genitalpapillen beschränkt ist[2]. Die Eier und Spermien werden zur äußerlichen Befruchtung reichlich ins Meerwasser entlassen[6]. Nach dem Ablaichen der Gameten und deren anschließender Befruchtung verläuft die Entwicklung von A. lixula über die für Echinoidea (Seeigel) typische Pluteuslarve. Sie zeichnet sich durch ihren bilateralsymmetrischen Aufbau aus, trägt 4–6 Armpaare und ihr Innenskelett besteht aus 4 symmetrischen und 1–2 unpaaren Teilen. Nach ungefähr 4–6 Wochen endet ihre planktotrophe Phase. Sie sinkt zu Grunde und durchläuft die Metamorphose zum benthischen, juvenilen 1 mm großen Seeigel. Diese Verwandlung dauert maximal 1 Stunde. A. lixula betreibt keine Brutpflege[4][2]. Der Erfolg ihrer Ansiedlung ist jedoch stark vom passenden Substrat abhängig. Der chemische Stimulus von Kalkalgen oder auch nackter Stein erleichtern die Niederlassung[17]. Nach Fénaux zeigen Untersuchungen des durch die Photoperiode gesteuerten gonadosomatischen Zyklus von A. lixula, dass dieser im Mittelmeer seinen Höhepunkt jährlich von Mai bis Juni erreicht, was mit dem Auftreten der Pluteuslarve im Plankton zwischen Juli und November korreliert[18]. Pedrotti konnte jedoch das Verschwinden der Larven von August bis September feststellen[19]. Da es sich hierbei um die wärmsten Monate und somit günstige Konditionen für die Entwicklung der Larve handelt, wird ein Zusammenhang dieser Beobachtung mit einer niedrigen Konzentration des Phytoplanktons während dieser Monate vermutet. Diese könnte aufgrund des Nahrungsmangels die Mortalitätsrate der Larven erhöhen. Alternativ wird ein größeres Auftreten planktivorer Fische vermutet, die die Larven des Schwarzen Seeigels frei[1]. In künstlichen Befruchtungsexperimenten können Individuen des Schwarzen Seeigels über eine KCL-Injektion zum Ablaichen gebracht werden. Die Aufzucht der Larven erweist sich als problemlos, weshalb sie als Modellorganismen der experimentellen Entwicklungsbiologie dienen.
Einfluss von Umweltbedingungen
BearbeitenTemperatur
BearbeitenA. lixula gilt als eine thermophile Art. Adulte Individuen sind ganzjährig auffindbar, Larven je nach Quelle vor allem im August[5]. Außerdem lässt sich nach heißen Sommern ein Populationsausbruch beobachten. Bei hohen Temperaturen zeigt diese Art eine höhere larvale Überlebensrate, welche durch die verkürzte Larvalperiode und der damit einhergehenden Verminderung von Prädation zu begründen ist[17]. Das Wachstum der Larven ist bei Temperaturen, die über der durchschnittlichen Meerestemperatur des Mittelmeers liegen, beschleunigt. Diese Beobachtung lässt vermuten, dass sich A. lixula im Mittelmeer in suboptimalen Bedingungen aufhält und dieses tatsächlich erst in jüngster zeit besiedelte. Dies würde allerdings auch bedeuten, dass eine Erwärmung des Mittelmeers im Zuge der globalen Erwärmung einen bisher limitierenden Faktor der A. lixula Populationen verschwinden lassen würde, wodurch sein negativer Effekt auf das küstennahe Ökosystem des Mittelmeeres künftig verstärkt werden könnte[1].
Salinität
BearbeitenIn Bezug auf vielerlei Arten wurde bereits festgestellt, dass eine Veränderung der Salinität ihres marinen Habitats deren Überleben, Reproduktion, Entwicklung oder auch Wachstum beeinflusst. Für A. lixula konnten in sogenannten Embryotoxizitätstests jedoch kein Einfluss einer Salinitätsveränderung auf die Gameten selbst oder deren Befruchtung nachgewiesen werden. So liegt der optimale Bereich der Salinität für die Entwicklung des Schwarzen Seeigels zwischen 29 und 35,5 Promille und deckt damit sogar einen höheren Toleranzbereich als die verwandte Art Paracentrotus lividus ab[8].
Nanopartikel
BearbeitenEs existieren zahlreiche Untersuchungen, die sich mit der Auswirkung von Nanopartikeln auf Umwelt und Organismen beschäftigen. Diese finden in Nanotechnologien verschiedener Bereiche wie der Medizin, Textilindustrie oder dem Ingenieurswesen Anwendung, wodurch sie häufig in die Umwelt gelangen und Einflüsse auf das Ökosystem nehmen können. Embryotoxizitätstests mit CuO-Nanopartikeln (Kupferoxid) zeigen eine konzentrationsabhängige Abnahme des Embryonenwachstums, Entwicklungsdefekte und Anomalitäten des Skeletts der Pluteuslarve von A. lixula. In kontaminiertem Meerwasser zeigt diese Art einen Schwund auf, da vor allem ihre frühen Entwicklungsstadien sensitiv auf den Kontakt mit CuO-Nanopartikeln reagieren. Aggregate dieser Nanopartikel werden vom Schwarzen Seeigel aufgenommen, sein cholinergisches und serotonergisches System zeigen Veränderungen auf und seine Neurotransmission werden beeinflusst[20].
Verwechslungsgefahr
BearbeitenDer Schwarze Seeigel wird aufgrund seiner äußerlichen Ähnlichkeit und Koexistenz häufig mit dem essbaren und wirtschaftlich stark genutzten Steinseeigel (Paracentrotus lividus) verwechselt. Fischer hielten diese ursprünglich für männliche und weibliche Individuen derselben Art. Darauf beruht auch deren jeweilige italienische Bezeichnung. So wurde der Schwarze Seeigel als riccio maschio und der Steinseeigel als riccio femina bezeichnet. Allerdings unterscheiden sich die lebenden Individuen bei genauer Betrachtung deutlich voneinander. Die peristomale Membran ist bei A. lixula deutlich größer und sichtbarer als bei P. lividus; sein stachelfreies Mundfeld nimmt also einen viel größeren Teil der Oralseite ein[7]. Außerdem hat der Steinseeigel kürzere und dichtere Stacheln und trägt im Gegensatz zum Schwarzen Seeigel auch auf der Aboralseite seines Körpers mit Saugnäpfen bestückte Ambulakralfüßchen[21]. Er neigt daher zur Maskierung, wozu A. lixula nicht befähigt ist. Der Steinseeigel ist gesteinsbohrend, weshalb man ihn oft in Wohnhöhlen am Felsen vorfinden und auch über dieses Merkmal sicher bestimmen kann[11]. Weiterhin ist die Laterne des Aristoteles von A. lixula größer, was deshalb, weil der herbivore P. lividus sich von nicht krustenbildenden, weicheren Organismen ernährt, als seine evolutive Anpassung an die Durophagie gesehen wird. Weiterhin ist die Gonadenmasse des Schwarzen Seeigels kleiner und der Steinseeigel zeichnet sich aufgrund der Verteilung seiner Ambulakralfüßchen am ganzen Körper durch eine sehr viel höhere Mobilität aus[10]. Lange Zeit wurde diesen beiden Arten aufgrund ihrer scheinbaren Vergesellschaftung dieselbe ökologische Nische und trophische Ebene zugeschrieben. Genauere Untersuchungen einer Interaktion müssen noch angestellt werden, jedoch zeigen unterschiedliche Substrat- und Nahrungsvorlieben, dass sich ihre Nische nicht vollständig überschneiden kann und die omnivore und teils karnivore Art A. lixula eine höhere trophische Ebene belegen muss. Dass diese Arten miteinander im Wettbewerb stehen, scheint daher unwahrscheinlich[1].
-
 Oralseite der Testa von A. lixula (l.) und P. lividus (r.)
Oralseite der Testa von A. lixula (l.) und P. lividus (r.) -
 Die Testa von A. lixula (l.) und P. lividus (r.) im Vergleich
Die Testa von A. lixula (l.) und P. lividus (r.) im Vergleich
_und_P._lividus_(r.).jpg)
_und_P._lividus_(r.)_im_Vergleich.jpg)
Bedeutung
BearbeitenÖkologisch
Bearbeiten.jpg)
A. lixula nimmt eine wichtige Rolle im Ökosystem des Litorals ein. Sein Weideverhalten führt zur Zerstörung von Algenwäldern sowie der Entstehung und Erhaltung von Kahlgebieten auf felsigen Substraten flacher mariner Ökosysteme. Diese Art kontrolliert die Dynamik, Struktur und den Aufbau von Populationen von Makroalgen[22]. Der Zusammenhang zwischen der Dichte dieser Art und der Erweiterung von Kahlgebieten ist eindeutig. Eine hohe Dichte des Schwarzen Seeigels kann Algen vollständig verschwinden lassen und dadurch die Produktivität, Struktur und Funktion des Ökosystems nachhaltig beeinflussen[23]. Darüber hinaus kann beobachtet werden, dass sich die Individuenanzahl von Napfschnecken der Gattung Patella bei Entfernung des Schwarzen Seeigels aus seinem natürlichen Habitat signifikant vermindert und in seiner Anwesenheit wieder ansteigt. Bulleri vermutet, dass die Anzahl dieser Vertreter der Napfschnecken von der Dichte der Makroalgen, die von A. lixula reguliert werden, abhängt[24]. Die Individuenanzahl der Rotalgengattung Peyssonnelia steigt jedoch bei Entfernung von A. lixula, was darauf schließen lässt, dass auch diese Vertreter der Rotalgen in Abhängigkeit zu ihm stehen. Insgesamt schreibt die regulierende Wirkung von Verteilung und Häufigkeit verschiedener Arten der benthischen Lebensgemeinschaft dem Schwarzen Seeigel eine signifikante Rolle im marinen Ökosystem zu. Die Verminderung seiner Prädatoren aufgrund der sich ändernden Umweltbedingungen und menschlichen Fischereiverhaltens, sowie die Tendenz zur Wassererwärmung, die den Fortpflanzungserfolg der thermophilen Art begünstigen, verstärken seine ökologische Rolle zunehmend. Aber auch diese Art benachteiligende Prozesse wie die mit der globalen Erwärmung einhergehende Versauerung der Meere sind zu beachten. Diese zeigen zwar keinen signifikanten Einfluss auf adulte Individuen, jedoch beeinträchtigen sie das Wachstum der Larven. Nichtsdestotrotz kann die ökologische Bedeutung dieser Art nur im Zusammenhang mit seiner Interaktion mit dem Steinseeigel vollständig erklärt werden und muss daher weiterhin erforscht werden[1].
Ökonomisch
Bearbeiten
Lange Zeit wurde der Schwarze Seeigel nicht wirtschaftlich genutzt. Allerdings sind seine Gonaden essbar und gelten im Mittelmeerraum als eine Delikatesse, die mit Zitronensaft beträufelt und roh verspeist werden. Der Schwund des häufiger verzehrten Steinseeigels bringt ihn nach und nach auch auf den Markt, wobei sein Geschmack als schwächer beschrieben wird. Die Individuen werden von Hand gesammelt und spezielle Seeigelöffner dienen als Werkzeug. Allerdings gilt zu beachten, dass die Gonaden dieser Art während der Fortpflanzungszeit giftig sein können und daher in diesem Zeitraum nicht verspeist werden sollten[14][7].
Bioindikator
BearbeitenAufgrund seiner Sensibilität auf verschiedene Umweltfaktoren und seiner Verfügbarkeit gilt A. lixula als wichtiger Bioindikator für das sogenannte Biomonitoring des marinen Ökosystems und besitzt auch dadurch eine große ökologische Signifikanz. Seine einfache Handhabung und die Möglichkeit zur Haltung in Aquarien begünstigen die Möglichkeiten der Umweltforschung anhand von Studien an dieser Art[20].
Medizinisch
BearbeitenDie intensive violette Färbung der Eier von A. lixula beruht auf der Einlagerung von Carotinoiden, die diese Art durch ihre Nahrung aufnimmt. Hierbei handelt es sich um Astaxanthin, ein Antioxidans, welches bei der Vorbeugung und Behandlung von Krebs, chronischen Entzündungserkrankungen, Erkrankungen des Stoffwechsels, der Leber, der Haut, der Augen oder des Darmes, Diabetes oder auch neurodegenerativen Erkrankungen unterstützen soll. Die Eier der kultivierten Individuen beinhalten eine 15-mal höhere Konzentration dieses Stoffes als die im natürlichen Lebensraum gesammelten Exemplare. Als künftige Quelle eines Stoffes, der pharmazeutische Signifikanz trägt, könnte sich die Medizin den Schwarze Seeigel in Kultur zu Nutzen machen ohne seine natürlichen Populationen zu erschöpfen[25].

Verletzung durch Arbacia lixula
BearbeitenEine Verletzung durch diese Art verursacht unangenehme und teils auch sehr schmerzhafte Verletzungen. Meist brechen die Stacheln in der Wunde ab und lösen sich zwar innerhalb weniger Tage auf, können jedoch durch eine Verschmutzung der Wunde starke Entzündungen verursachen. Entgegen der Annahme vieler Touristen sind die Stacheln des Schwarzen Seeigels allerdings nicht giftig[3].
Zusätzliches Bildmaterial
Bearbeiten-
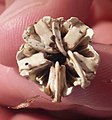 Hinteransicht der in die Testa eingelagerten Laterne
Hinteransicht der in die Testa eingelagerten Laterne -
 Seitenansicht der Laterne mit ihren pfeilförmigen Pyramiden
Seitenansicht der Laterne mit ihren pfeilförmigen Pyramiden -
 Ambulakralfüßchen des Violetten Seeigels (Sphaerechinus granularis) mit Saugnäpfen
Ambulakralfüßchen des Violetten Seeigels (Sphaerechinus granularis) mit Saugnäpfen -
 Aboralseite von A. lixula
Aboralseite von A. lixula


_mit_Saugn%C3%A4pfen.jpg)

Einzelnachweise
Bearbeiten- ↑ a b c d e f g h i Wangensteen, O. S. (2013). Biology and phylogeography of the black sea urchin Arbacia lixula (Echinoidea: Arbacioida) (Doctoral dissertation, Ph. D. Thesis, Universitat de Barcelona, Spain).
- ↑ a b c d e f g Westheide, W., Rieger, R., Dohle, W., Emschermann, P., Goetting, K. J., Goldschmid, A.,... & Huelsmann, N. (2013). Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere. Springer Spektrum, Heidelberg.
- ↑ a b c d e f DeHaas, W., Knorr, F. (1999). Was lebt da im Meer an Europas Küsten? Müller Rüschlikon, Stuttgart.
- ↑ a b c Riedl, R. (1983). Fauna und Flora des Mittelmeeres. P. Parey.
- ↑ a b Tortonese, E., 1965. Echinodermata. Fauna d’Italia vol. VI. Calderini, Bologna.
- ↑ a b c Hickman, C. P. (2008). Zoologie. Pearson Deutschland GmbH, Hallbergmoos.
- ↑ a b c d Tortonese, E., & Vadon, C. (1987). Oursins et Holothuries (Echinodermes). Fiches d’identification des especes pour les besoins de la peche, 1, 715-739.
- ↑ a b c Carballeira, C., Martín-Díaz, L., & DelValls, T. A. (2011). Influence of salinity onfertilization and larval development toxicity tests with two species of sea urchin. Marine environmental research, 72(4), 196-203.
- ↑ Bonaviri, C., Vega Fernández, T., Fanelli, G., Badalamenti, F., Gianguzza, P., 2011. Leading role of sea urchin Arbacia lixula in maintaining barren state in southwestern Mediterranean. Marine Biology 158, 2505–2513.
- ↑ a b c d e Agnetta, D., Bonaviri, C., Badalamenti, F., Scianna, C., Vizzini, S., Gianguzza, P., 2013. Functional traits of two co-occurring sea urchins across a barren/forest patch system. J. Sea Res. 76, 170-177.
- ↑ a b Göthel, H. (1997). Niedere Tiere Farbatlas Mittelmeerfauna: Niedere Tiere und Fische. Ulmer, Stuttgart.
- ↑ Agnetta, D., Badalamenti, F., Ceccherelli, G., Di Trapani, F., Bonaviri, C., & Gianguzza, P. (2015). Role of two co-occurring Mediterranean sea urchins in the formation of barren from Cystoseira canopy. Estuarine, coastal and shelf science, 152, 73-77.
- ↑ Benedetti-Cecchi, L., Bulleri, F., Cinelli, F., 1998. Density dependent foraging of sea urchins in shallow subtidal reefs on the west coast of Italy (western Mediterranean). Mar. Ecol. Prog. Ser. 163, 203–211.
- ↑ a b Hofrichter, R. (2004). Das Mittelmeer: Fauna, Flora, Ökologie-Bestimmungsführer, II/1. Spektrum Akademischer Verlag, Heidelberg.
- ↑ Guidetti, P., Mori M (2005) Morpho-functional defences of Mediterranean sea urchins, Paracentrotus lividus and Arbacia lixula, against fish predators. Mar Biol 147: 797– 802.
- ↑ Guidetti, P., (2004) Consumers of sea urchins (Paracentrotus lividus and Arbacia lixula) in shallow Mediterranean rocky reefs. Helgol Mar Res 58:110–116.
- ↑ a b Privitera, D., Noli, M., Falugi, C., & Chiantore, M. (2011). Benthic assemblages and temperature effects on Paracentrotus lividus and Arbacia lixula larvae and settlement. Journal of Experimental Marine Biology and Ecology, 407(1), 6-11.
- ↑ Fénaux, L., 1968. Maturation des gonades et cycle saisonnier des larves chez A. lixula, P. lividus et P. microtuberculatus à Villefranche-Sur-Mer. Vie et Milieu Série A Biologie Marine 19, 1–52.
- ↑ Pedrotti, M.L., 1993. Spatial and temporal distribution and recruitment of echinoderm larvae in the Ligurian Sea. Journal of the Marine Biological Association of the United Kingdom 73, 513–530.
- ↑ a b Maisano, M., Cappello, T., Catanese, E., Vitale, V., Natalotto, A., Giannetto, A. & Fasulo, S. (2015). Developmental abnormalities and neurotoxicological effects of CuO NPs on the black sea urchin Arbacia lixula by embryotoxicity assay. Marine environmental research, 111, 121-127.
- ↑ Santos, R., Flammang, P., 2005. Morphometry and mechanical design of tube foot stems in sea urchins: a comparative study. J. Exp. Mar. Biol. Ecol. 315, 211-223.
- ↑ Hereu, B., 2006. Depletion of palatable algae by sea urchins and fishes in a Mediterranean subtidal community. Mar. Ecol. Progr. Ser. 313, 95–103.
- ↑ Sala, E., 2004. The past and present topology and structure of Mediterranean subtidal rocky-shore food webs. Ecosystems 7, 333–340.
- ↑ Bulleri, F., Benedetti-Cecchi, L., & Cinelli, F. (1999). Grazing by the sea urchins Arbacia lixula L. and Paracentrotus lividus Lam. in the Northwest Mediterranean. Journal of Experimental Marine Biology and Ecology, 241(1), 81–95.
- ↑ Cirino, P., Brunet, C., Ciaravolo, M., Galasso, C., Musco, L., Vega Fernández, T., & Toscano, A. (2017). The sea urchin Arbacia lixula: a novel natural source of astaxanthin. Marine drugs, 15(6), 187.