Eukaryoten oder Eukaryonten (Eukaryota) (von altgriechisch εὖ eu = ‚richtig‘, ‚gut‘ und κάρυον karyon = ‚Nuss‘, ‚Kern‘) sind eine Domäne der Lebewesen, deren Zellen (Eucyten) einen echten Kern und eine reiche Kompartimentierung haben[1] (Tiere, Pflanzen und Pilze). Hierin unterscheiden sie sich von den beiden übrigen Domänen im System der Lebewesen, den prokaryotischen Bakterien und Archaeen (letztere früher auch Urbakterien genannt), beide mit procytischen Zellen.
| Eukaryoten | ||||
|---|---|---|---|---|

Verschiedene Eukaryoten | ||||
| Systematik | ||||
| ||||
| Wissenschaftlicher Name | ||||
| Eukaryota | ||||
| Chatton, 1925 |



Merkmale
BearbeitenDie Zellen der Eukaryoten haben meistens einen Durchmesser von 10 bis 30 µm.[2] Sie sind in der Regel wesentlich größer als die von Prokaryoten, ihr Volumen beträgt etwa das 100- bis 10.000-Fache. Für ein reibungsloses Funktionieren der zellulären Abläufe über größere Entfernungen innerhalb der Zelle sind ein höherer Organisationsgrad und eine Aufteilung des Zellraums in Kompartimente (abgegrenzte Räume) sowie Transport zwischen diesen Kompartimenten günstig. Aus diesem Grund sind eukaryotische Zellen mittels Zellorganellen strukturiert, welche wie die Organe eines Körpers verschiedene Funktionen ausüben. Das namensgebende Organell ist der Zellkern mit dem Hauptanteil des genetischen Materials der eukaryotischen Zellen. Weitere Gene kommen je nach Art in Mitochondrien (Organellen, die durch chemische Reaktionen Energie zur Verfügung stellen), in Einzelfällen auch in den ihnen verwandten Hydrogenosomen sowie in fast allen Plastiden (unter anderem in den Photosynthese betreibenden Chloroplasten) vor. Dem intrazellulären Transport dienen die Organellen des Endomembransystems.
Struktur und Form wird den eukaryotischen Zellen durch das Cytoskelett verliehen, das auch der Fortbewegung dient. Es ist aus Mikrotubuli, Intermediärfilamenten und Mikrofilamenten aufgebaut. Einige Eukaryoten, beispielsweise Pflanzen und Pilze, haben auch Zellwände, die die Zellen außerhalb der Cytoplasmamembran einschließen und ihre Form bestimmen.
Eine weitere Besonderheit der Eukaryoten liegt in der Proteinbiosynthese: Anders als Prokaryoten sind Eukaryoten in der Lage, mittels derselben DNA-Information durch alternatives Spleißen unterschiedliche Proteine herzustellen.
Mehrzelligkeit
BearbeitenEukaryoten können Einzeller oder mehrzellige Lebewesen sein. Diese bestehen aus einer größeren Zahl von Zellen mit gemeinsamem Stoffwechsel, wobei spezielle Zelltypen bestimmte Aufgaben übernehmen. Die meisten bekannten Mehrzeller sind Eukaryoten, darunter die Pflanzen, Tiere und mehrzelligen Pilze.
Systematik
Bearbeiten

Alastair Simpson, Dalhousie University, 2020
Der Name verweist auf das Vorhandensein eines Zellkerns (Eukaryota/Eukaryonta/Eucarya/Eukarya; zu altgriechisch εὖ eû ‚gut, echt‘ und κάρυον káryon ‚Nuss‘).
In der biologischen Systematik bilden die Eukaryoten eine der drei Domänen, also der höchsten Kategorien zur Klassifizierung von Lebewesen. Die aktuelle Systematik der Eukaryoten wurde von Adl et al. 2019 aufgestellt. Sie gliedert die Eukaryoten wie folgt:[3]
- Amorphea
- super-group Amoebozoa, einzellige Organismen mit im Regelfall amöboider Gestalt, inkl. Schleimpilze und manche Amöben
- super-group Obazoa, bestehend aus Opisthokonta, inkl. Tiere und Pilze, Breviatea und Apusomonadida
- Diaphoretickes
- super-group Archaeplastida, inkl. Pflanzen
- super-group Sar, inkl. Stramenopile (auch als Stramenopiles, Chromista oder Heterokonta bezeichnet)
- einige Taxa unsicherer Stellung, darunter Cryptista (inkl. Cryptophyceae), Haptista (inkl. Haptophyta), Picozoa (neulich betrachtet als Schwestergruppe der Rotalgen[4]), Rappemonads, Telonemia

Daneben existieren zahlreiche Taxa mit unsicherer Stellung (incertae sedis), darunter unter anderem:[5]
Weitere Taxa incertae sedis sind die Formgruppe der „Excavates“, früher betrachtet als super-group Excavata, einzellige Organismen, die mehrheitlich begeißelt sind; darunter:
Systematik nach Cavalier-Smith
Bearbeiten
Thomas Cavalier-Smith gliedert die Eukaryoten wie folgt:[7][8][9][8][10][11][12][13]
| Eukaryota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
*) syn. Ancyromonadida
Entwicklungsgeschichte
Bearbeiten

Mikrofossilien
BearbeitenDie ältesten makroskopischen, mehrzelligen, möglicherweise eukaryotischen Fossilien sind 1,5 Milliarden Jahre alt.[16] Es ist nicht bekannt, ob die hypothetischen evolutionären Vorläufer der Eukaryoten – die sogenannten Urkaryoten[17] – bereits Organellen besaßen (diese wären aufgrund des nachteiligen Oberflächen-Volumen-Verhältnisses nötig gewesen) oder sie erst im Laufe ihrer Evolution hin zu den eigentlichen Eukaryoten erwarben. Die bekannteste Theorie zur Entstehung der Organellen ist die Endosymbiontentheorie, die besagt, dass Mitochondrien und Chloroplasten auf Bakterien zurückgehen, die von den zellbiologisch „primitiven“, räuberisch lebenden Urkaryoten ursprünglich als Nahrung aufgenommen und von denen einige durch Zufall nicht verdaut wurden, sondern eine Symbiose mit dem Urkaryoten eingingen.[18][19]
LECA
BearbeitenDer letzte gemeinsame Vorfahr aller heutigen Eukaryoten – in der meist englischen Fachliteratur auch als Last Eukaryotic Common Ancestor (LECA), bezeichnet[20] – sollte sowohl einen abgegrenzten Zellkern als auch Mitochondrien besessen haben, da alle bisher untersuchten Eukaryoten Mitochondrien, mitochondrienartige Organellen oder zumindest Kern-DNA von mitochondrialem Ursprung (übertragen durch lateralen Gentransfer) besitzen. Zwar führte Thomas Cavalier-Smith für rezente amitochondriale Einzeller mit Zellkern, die er für evolutionäre Relikte aus der Zeit vor LECA hielt, die Bezeichnung Archezoa ein,[21] jedoch mehrten sich nachfolgend Hinweise darauf, dass es sich bei den Archezoen um sekundär amitochondriale echte Eukaryoten handelt,[22] das heißt um Abkömmlinge von LECA.
Mitochondrienerwerb
BearbeitenAls Alternative zur Urkaryoten-Hypothese wurde 1998 von Bill Martin und Miklós Müller mit der sog. Wasserstoff-Hypothese (en. hydrogen hypothesis) der Erwerb von Mitochondrien durch Archaeen vorgeschlagen.[23][24] Es hat sich nämlich gezeigt, dass die DNA rezenter (heutiger) Asgard-Archaeen enger mit der DNA in den eukaryotischen Zellkernen verwandt ist, als mit der DNA anderer Archaeen. Die Asgard-Archaeen leben in anoxischen Sedimenten der Ozeane und können mit Bakterien in Symbiose leben. Dies entspricht den Voraussetzungen der Wasserstoff-Hypothese, wonach die ersten eukaryotischen Zellen (Eucyten) durch Erwerb der Mitochondrien unter anoxischen Bedingungen entstanden sind.[25]
Nach 12 Jahren Forschungsarbeit berichteten Mikrobiologen 2019, erfolgreich ein sich nur sehr langsam vermehrendes Lokiarchaeon aus Tiefseeschlamm isoliert und kultiviert zu haben.[15][26][27] Candidatus Prometheoarchaeum syntrophicum ist ein, nach Vergleich seines Genoms nahe mit den Eukaryoten verwandtes, extrem langsam wachsendes heterotrophes Archaeon, das Energie aus dem Abbau von Peptiden und Aminosäuren gewinnt und dabei Wasserstoff produziert und abgibt. Das Archaeon gedeiht nur in obligater Syntrophie mit Wasserstoff-metabolierenden Organismen, einer davon ein sulfatreduzierendes Delta-Proeobakterium der Desulfovibrionales. Die Autoren nehmen an, dass treibende Kraft der Endosymbiose, die zu den Eukaryoten führte, ein zweites, in ihrem Modellsystem nicht vorhandenes Bakterium, das künftige Mitochondrium, gewesen wäre, dessen Aufgabe zunächst primär die Beseitigung von für das obligat anaerobe Archaeon giftigen Sauerstoffs gewesen sei. Die Rolle des Mitochondriums im Energiestoffwechsel wäre demnach erst sekundär entstanden. Die Autoren halten eine Schlüsselrolle des Wasserstoff-Metabolismus selbst, wie in alternativen Szenarien der Endosymbiontentheorie[28] vorgeschlagen, für unplausibel. Dementsprechend wäre das Wasserstoff-verwertende Bakterium selbst nicht zum Endosymbionten geworden. Die Hypothese setzt einen frühen Ursprung des Mitochondriums in einer noch primitiven, Archaeen-artigen Wirtszelle voraus und unterscheidet sich darin von alternativen Szenarien wie der „Archezoa“-Hypothese von Thomas Cavalier-Smith, nach denen ein fast fertiger Pro-Eukaryot erst spät ein Mitochondrium erworben hätte. Dementsprechend wäre ein „Einfangen“ des künftigen Mitochondriums über Phagocytose unplausibel, da es an den nötigen internen Strukturen wie einem Cytoskelett gefehlt hätte. Interessanterweise verfügt Prometheoarchaeum aber über tentakel-artige Zellfortsätze, mit denen ein ähnlicher Organismus im Archaikum den hypothetischen Endosymbionten alternativ quasi umschlungen haben könnte.
Im Jahr 2023 identifizierten Laura Eme, Daniel Tamarit et al. auf Basis einer vergleichenden Analyse von Asgard-Genomen die Ordnung Hodarchaeales innerhalb der Heimdallarchaeia als Schwesterklade der Eukaryoten unter den zu diesem Zeitpunkt bekannten Archaeen.[29]
Asgardviren
BearbeitenDieser Erwerb könnte vor oder gleichzeitig mit dem des Zellkerns stattgefunden haben und könnte eventuell viralen Ursprung sein (Virale Eukaryogenese, „viral eukaryogenesis“, ein Teil der „Out-of-Virus-Hypothese“).[30] Als Kandidaten werden DNA-Viren beispielsweise unter Riesenviren der Nucleocytoviricota (alias Nucleocytoplasmic large DNA viruses, NCLDVs), wie etwa die vorgeschlagenen „Medusaviridae“ gehandelt.[31] Aber auch der „Pseudomonas Phage 201phi2-1“[32][33] aus der Gattung Phikzvirus, ein Riesen-Bakteriophage (Caudoviricetes, Morphotyp Myoviren), bildet bei der Infektion einer Bakterienzelle eine kernähnliche Struktur, die Proteine abhängig von ihrer Funktion trennt.[34][35] Diese kernartige Struktur und ihre Schlüsseleigenschaften wurden auch in verwandten Phagen gefunden.[36] Auftrieb bekam diese Hypothese 2021/22 durch die Entdeckung verschiedener Viren der Asgard-Archaeen (kurz Asgardviren genannt). Diese zeigen Merkmale prokaryotischer Viren (z. B. mit Kopf-Schwanz-Aufbau – Klasse Caudoviricetes, oder ellipsoid- bis spindelförmiger Aufbau – Familien Ovaliviridae, Bicaudaviridae und Fuselloviridae) als auch eukaryotischer Viren (wie der NCLDVs).[37][38][39][40]
Reihenfolge
BearbeitenEine im Herbst 2020 veröffentlichte Studie legt anhand von umfangreichen Genomanalysen tatsächlich nahe, dass – obwohl bisher noch keine primär amitochondrialen Eukaryoten gefunden wurden – die Vorfahren der Eukaryonten zuerst ihr komplexes Genom mit den zugehörigen Strukturen, und danach die Mitochondrien (oder Vorläufer davon) erworben haben.[41] Eine Zusammenfassung dieser Diskussion auf dem Stand von 2019 findet sich bei Traci Watson;[42] zu den unterschiedlichen Standpunkten siehe Guglielmini et al. (2019)[43] und Koonin und Yutin (2018).[44]
Die Entstehung der Chloroplasten der Photosynthese betreibenden Eukaryoten durch Endosymbiose stellt einen zeitlich späteren Vorgang dar. Zunächst wurden Cyanobakterien von nicht-phototrophen Eukaryoten aufgenommen, was möglicherweise in dieser Form (primäre Endosymbiose) nur einmal geschah. Die anderen Plastiden (Leukoplasten etc.) leiten sich von diesen ab.
Später entstanden komplexe Plastiden (mit mehr als doppelter Membran und ggf. Nukleosom) durch eine sekundäre Endosymbiose, d. h. (weitere) nicht-phototrophe Eukaryoten nahmen (aus der primären Endosymbiose entstandene) phototrophe Eukaryoten durch Endosymbiose auf (so etwa bei den Apicomplexa[45]).
Forschungsgeschichte
BearbeitenDie Einteilung von Lebewesen in Prokaryoten und Eukaryoten wurde erstmals von Edouard Chatton für Protisten deutlich herausgestellt und 1925 veröffentlicht.[46][47]
Diese Einteilung trat zunächst mit der Einführung des Drei-Domänen-Systems durch Carl Woese 1977 in den Hintergrund,[48][49] eines Ansatzes, der das zelluläre Leben in die drei Domänen Bakterien, Archaeen und Eukaryoten aufteilt.
Im Jahr 1984 wurde von James A. Lake und Kollegen die Eozyten-Hypothese vorgeschlagen: Man hatte entdeckt, dass bei der Form der Ribosomen in der Archaeen-Gruppe der Crenarchaeota (mit der ursprünglichen Bezeichnung Eozyten) und der Eukaryoten eine erstaunliche Ähnlichkeit besteht, dass aber die Form der Ribosomen sowohl bei den Bakterien als auch bei den Euryarchaeota, einer anderen Archaeen-Gruppe, deutlich davon abweicht. Es wurde daher vermutet, dass die Eukaryoten aus den Crenarchaeota hervorgegangen sind.[50][51]
Trotz weiterer Hinweise in den 1980er Jahren bekam die Eozyten-Hypothese erst in den 2000er Jahren mit dem Fortschritt der Genom-Analyse neuen Auftrieb. In einer ganzen Reihe von Archaeen wurden Gene entdeckt, die man in ähnlicher Form auch in Eukaryoten findet. Entsprechend den Ergebnissen einiger Studien wurde in einer Abwandlung statt der Crenarchaeota die diesen nahestehende Gruppe der Thaumarchaeota als Ursprung der Eukaryoten vorgeschlagen. (Die Crenarchaeota und Thaumarchaeota werden zusammen mit einigen anderen Archaeen-Gruppen in der Supergruppe „TACK“ zusammengefasst).[50][52][53][54][55]
Mit dem Aufkommen der Metagenomanalyse ließen sich seit 2015 in Proben aus der Nähe von Hydrothermalquellen Kandidaten für Archaeen-Gruppen identifizieren, die den Eukaryoten noch viel näher stehen müssen, als alle zuvor betrachteten Gruppen. Die erste Gruppe war die der „Lokiarchaeota“, gefunden bei einem hydrothermalen Vent namens „Lokis Schloss“ im Arktischen Ozean zwischen Mohns- und Knipovitsch-Kamm.[56] Da diese Erkenntnisse lediglich aus einer Metagenom-Analyse stammen und man die betreffenden Mikroben noch bis auf weiteres nicht kultivieren kann, kommt allen fraglichen Taxa lediglich ein 'Kandidatenstatus' zu, gekennzeichnet durch die Anführungszeichen.
Die „Lokiarchaeota“ werden aufgrund von Ähnlichkeit im Genom mit einigen ebenfalls in diesem Zusammenhang jüngst vorgeschlagenen Gruppen („Thorarchaeota“, „Odinarchaeota“ und „Heimdallarchaeota“) zu einer Kandidaten-Gruppe „Asgard“ zusammengefasst, die unter den Archaeen den Eukaryoten somit am nächsten steht und ein Schwester-Taxon zur Supergruppe „TACK“ darstellt.[57][58][59][60]
Um der engen Verwandtschaft der Archaeen mit den Eukaryoten Rechnung zu tragen, hat Thomas Cavalier-Smith sie bereits 2002 in ein gemeinsames Taxon Neomura gestellt, das damit eine Schwestergruppe zu den Bakterien darstellt – allerdings hatte er ursprünglich angenommen, dass die Trennung von Eukaryoten und Archaeen frühzeitig erfolgte, nicht dass die Eukaryoten aus der „Mitte“ der Archaeen entspringen.[61]
Diese neue Einteilung wird unterstützt durch neuere Erkenntnisse, wonach die Verwendung des DNA-Genoms als Träger der Erbinformationen bei Bakterien einerseits und Archaeen und Eukaryoten andererseits unterschiedlichen (möglicherweise viralen: anderer Teil der „Out-of-Virus-Hypothese“) Ursprungs zu sein scheint. Der letzte gemeinsame Vorfahr aller heute bekannten Lebewesen wäre dann ein archaischer zellulärer Organismus der RNA-Welt (mit Ribosomen, d. h. Protein-Synthese) gewesen.[30]
Weitere Unterstützung bekommt die Theorie durch Untersuchung des Aufbaus der membranständigen F- und V-/A-Typ-ATPasen. Die F-Typ-ATPasen der Mitochondrien und Chloroplasten sind mit denen der Bakterien homolog (wie nach der Endosymbiosetheorie zu erwarten). Die V-Typ-ATPasen an Zytoplasma-Membranen (Vakuolen) der Eukaryoten sind zu denen der Archaeen homolog, was eine Abstammung der Eukaryoten aus einem Zweig der Archaeen stützt. Vereinzelte Ausnahmen (F-Typ-ATPasen bei einigen Archaeen-Species[62] und V-Typ-ATPasen bei einigen Bakteriengruppen) werden auf horizontalen Gentransfer zurückgeführt.[63] Zentrale Untereinheiten der ATPasen sind über alle Lebewesen homolog, was für einen LUCA mit einer zumindest primitiven Membranhülle spricht.[64][65]
Einzelnachweise
Bearbeiten- ↑ Eukaryoten. In: Lexikon der Biologie. Spektrum Akademischer Verlag. Heidelberg, 1999, abgerufen am 1. Oktober 2016.
- ↑ Gerald Karp, Molekulare Zellbiologie, 2005, S. 25.
- ↑ Sina M. Adl, David Bass, Christopher E. Lane, Julius Lukeš, Conrad L. Schoch, Alexey Smirnov, Sabine Agatha, Cedric Berney, Matthew W. Brown, Fabien Burki, Paco Cárdenas, Ivan Čepička, Lyudmila Chistyakova, Javier del Campo, Micah Dunthorn, Bente Edvardsen, Yana Eglit, Laure Guillou, Vladimír Hampl, Aaron A. Heiss, Mona Hoppenrath, Timothy Y. James, Anna Karnkowska, Sergey Karpov, Eunsoo Kim, Martin Kolisko, Alexander Kudryavtsev, Daniel J. G. Lahr, Enrique Lara, Line le Gall, Denis H. Lynn, David G. Mann, Ramon Massana, Edward A. D. Mitchell, Christine Morrow, Jong Soo Park, Jan W. Pawlowski, Martha J. Powell, Daniel J. Richter, Sonja Rueckert, Laura Shadwick, Satoshi Shimano, Frederick W. Spiegel, Guifré Torruella, Noha Youssef, Vasily Zlatogursky, Qianqian Zhang: Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. In: The Journal of Eukaryotic Microbiology. Band 66, Nr. 1, 26. September 2018; doi:10.1111/jeu.12691, ISSN 1550-7408, ISSN 1066-5234, S. 4–119 (englisch; PDF-Datei, 1,49 MB, in der Wiley Online Library von John Wiley & Sons, Inc.).
- ↑ Max E. Schön, Vasily V. Zlatogursky, Rohan P. Singh, Camille Poirier, Susanne Wilken, Varsha Mathur, Jürgen F. H. Strassert, Jarone Pinhassi, Alexandra Z. Worden, Patrick J. Keeling, Thijs J. G. Ettema, Jeremy G. Wideman, Fabien Burki: Single cell genomics reveals plastid-lacking Picozoa are close relatives of red algae. In: Nature Communications. Band 12: 6651, 17. November 2021, doi:10.1038/s41467-021-26918-0, PMID 34789758, PMC 8599508 (freier Volltext), ISSN 2041-1723 (englisch)
- ↑
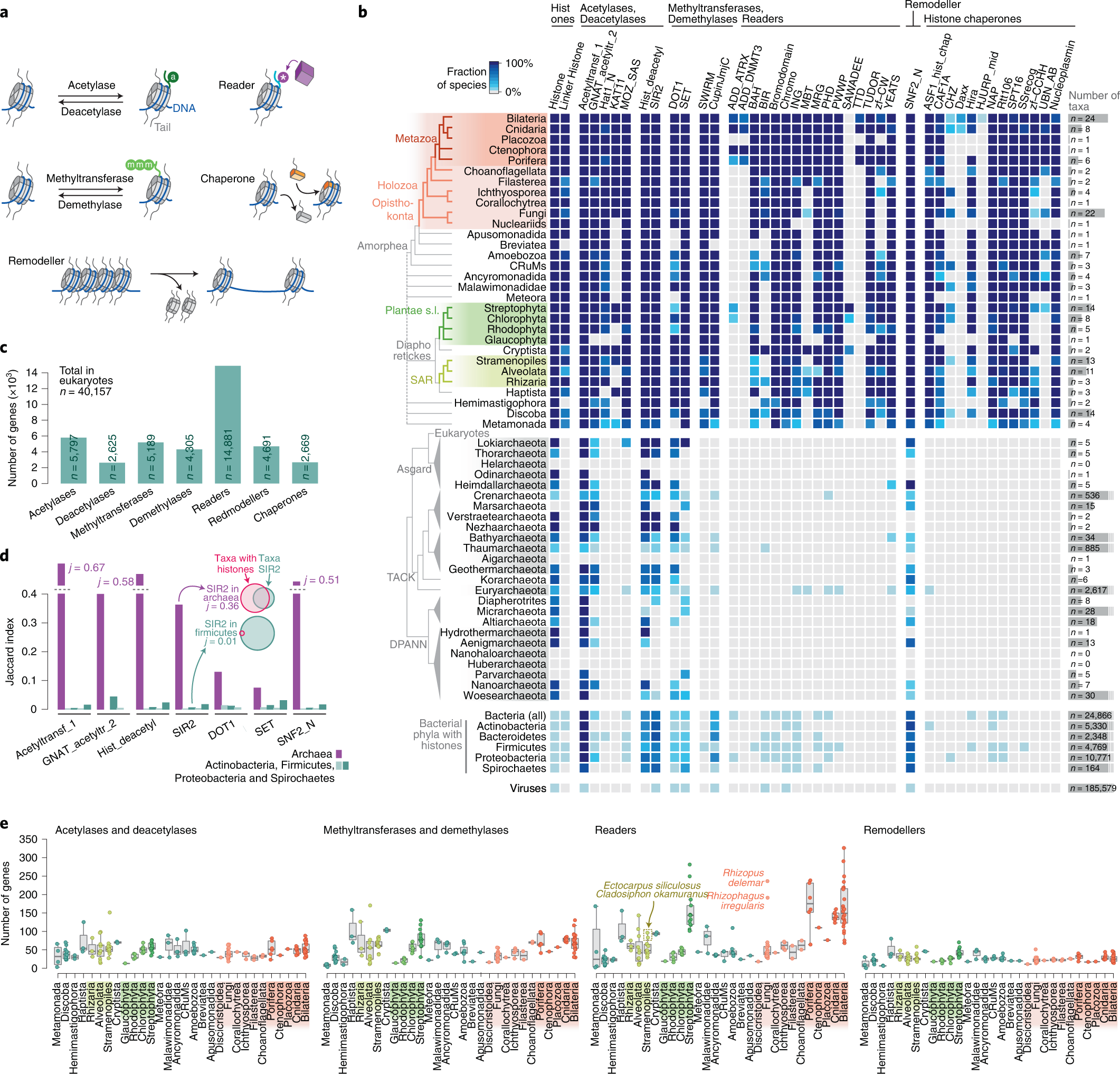
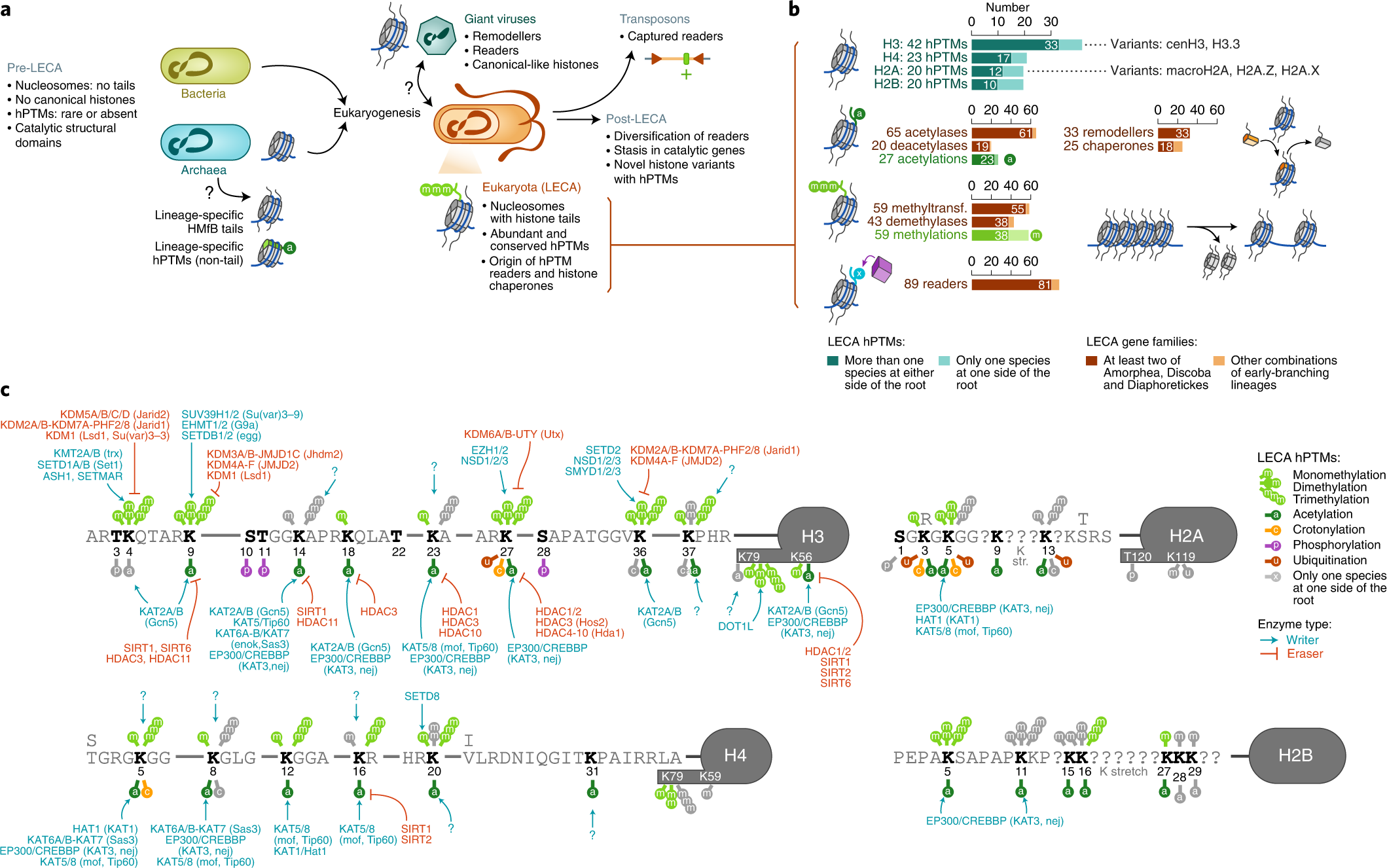
Xavier Grau-Bové, Cristina Navarrete, Cristina Chiva, Thomas Pribasnig, Meritxell Antó, Guifré Torruella, Luis Javier Galindo, Bernd Franz Lang, David Moreira, Purificación López-Garcia, Iñaki Ruiz-Trillo, Christa Schleper, Eduard Sabidó, Arnau Sebé-Pedrós: A phylogenetic and proteomic reconstruction of eukaryotic chromatin evolution. In: Nature Ecology & Evolution, Band 6, S. 1007–1023, 9. Juni 2022; doi:10.1038/s41559-022-01771-6, PMID 35680998 PMC 7613034 (freier Volltext). Dazu:
- Fig. 1: Diversity of post-translational modifications in eukaryotic canonical and variant histones.
- Fig. 3: Taxonomic distribution of chromatin-associated gene classes.
- Fig. 6: Chromatin evolution and eukaryogenesis.
- Xavier Grau-Bové: Repository: Chromatin evolution. Auf: GitHub.
- Chromatin First Evolved in Ancient Microbes 1-2 Billion Years Ago, New Research Suggests. Auf: sci-news vom 13. Juni 2022.
- Chromatin originated in ancient microbes one to two billion years ago. Genomic and proteomic analysis reveals that the regulatory role of chromatin is a eukaryotic innovation. Auf: ScienceDaily vom 9. Juni 2022. Quelle: Center for Genomic Regulation (CRG)
- Chromatin originated in ancient microbes one to two billion years ago. Center for Genomic Regulation (CRG) vom 9. Juni 2022.
- ↑ Klaus Hausmann, Markus Weitere, Matthias Wolf, Hartmut Arndt: Meteora sporadica gen. nov. et sp. nov. (Protista incertae sedis) – an extraordinary free-living protist from the Mediterranean deep sea. In: European Journal of Protistology, Band 38, Nr. 2, 2002, S. 171-177; doi:10.1078/0932-4739-00872, Epub 3. November 2004.
- ↑ Romain Derelle, Guifré Torruella, Vladimír Klimeš, Henner Brinkmann, Eunsoo Kim, Čestmír Vlček, B. Franz Lang, Marek Eliáš: Bacterial proteins pinpoint a single eukaryotic root. In: Proceedings of the National Academy of Sciences. 112. Jahrgang, Nr. 7, 17. Februar 2015, ISSN 0027-8424, S. E693–E699, doi:10.1073/pnas.1420657112, PMID 25646484, PMC 4343179 (freier Volltext) – (englisch, pnas.org).
- ↑ a b Thomas Cavalier-Smith, Ema E.Chao, Elizabeth A.Snell, Cédric Berney, Anna Maria Fiore-Donno, Rhodri Lewis: Multigene eukaryote phylogeny reveals the likely protozoan ancestors of opisthokonts (animals, fungi, choanozoans) and Amoebozoa. In: Molecular Phylogenetics & Evolution. 81. Jahrgang, Dezember 2014, S. 71–8 5, doi:10.1016/j.ympev.2014.08.012 (sciencedirect.com).
- ↑ Thomas Cavalier-Smith: Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree. In: Biology Letters. 6. Jahrgang, Nr. 3, 23. Juni 2010, ISSN 1744-9561, S. 342–345, doi:10.1098/rsbl.2009.0948, PMID 20031978, PMC 2880060 (freier Volltext) – (englisch, royalsocietypublishing.org).
- ↑ Ding He, Omar Fiz-Palacios, Cheng-Jie Fu, Johanna Fehling, Chun-Chieh Tsai, Sandra L. Baldauf: An Alternative Root for the Eukaryote Tree of Life. In: Current Biology. 24. Jahrgang, Nr. 4, S. 465–470, doi:10.1016/j.cub.2014.01.036.
- ↑ Laura A. Hug, Brett J. Baker, Karthik Anantharaman, Christopher T. Brown, Alexander J. Probst, Cindy J. Castelle, Cristina N. Butterfield, Alex W. Hernsdorf, Yuki Amano: A new view of the tree of life. In: Nature Microbiology. 1. Jahrgang, Nr. 5, 11. April 2016, ISSN 2058-5276, doi:10.1038/nmicrobiol.2016.48 (englisch, nature.com).
- ↑ Thomas Cavalier-Smith: Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. In: Protoplasma. 5. September 2017, ISSN 0033-183X, S. 1–61, doi:10.1007/s00709-017-1147-3 (englisch).
- ↑ Thomas Cavalier-Smith: Euglenoid pellicle morphogenesis and evolution in light of comparative ultrastructure and trypanosomatid biology: semi-conservative microtubule/strip duplication, strip shaping and transformation. In: European Journal of Protistology. Oktober 2017, doi:10.1016/j.ejop.2017.09.002 (elsevier.com).
- ↑ Eugene V. Koonin: Energetics and population genetics at the root of eukaryotic cellular and genomic complexity. In: Proceedings of the National Academy of Sciences. 112. Jahrgang, Nr. 52, 29. Dezember 2015, S. 15777–15778, doi:10.1073/pnas.1520869112 (pnas.org).
- ↑ a b Hiroyuki Imachi, Masaru K. Nobu, Nozomi Nakahara, Yuki Morono, Miyuki Ogawara, Yoshihiro Takaki, Yoshinori Takano, Katsuyuki Uematsu, Tetsuro Ikuta, Motoo Ito, Yohei Matsui, Masayuki Miyazaki, Kazuyoshi Murata, Yumi Saito, Sanae Sakai, Chihong Song, Eiji Tasumi, Yuko Yamanaka, Takashi Yamaguchi, Yoichi Kamagata, Hideyuki Tamaki & Ken Takai: Isolation of an archaeon at the prokaryote–eukaryote interface. In: Nature. Band 577, 2020, S. 519–525, doi:10.1038/s41586-019-1916-6.
- ↑ Eukaryoten: Eine neue Zeittafel der Evolution. Newsroom, Max-Planck-Gesellschaft, 24. Mai 2015.
- ↑ C. R. Woese, G. E. Fox: Phylogenetic structure of the prokaryotic domain: the primary kingdoms. In: Proceedings of the National Academy of Sciences. Band 74, Nummer 11, November 1977, S. 5088–5090, PMID 270744, PMC 432104 (freier Volltext).
- ↑ Lynn Margulis, Dorion Sagan: Leben: Vom Ursprung zur Vielfalt. Spektrum Akademischer Verlag, Heidelberg, Berlin 1997, ISBN 3-8274-0524-6 (Übersetzung der englischsprachigen Originalausgabe von 1995).
- ↑ Lynn Margulis: Die andere Evolution. Spektrum Akademischer Verlag, Heidelberg, Berlin 1999, ISBN 3-8274-0294-8 (Übersetzung der englischsprachigen Originalausgabe von 1998).
- ↑ Eugene V. Koonin: The origin and early evolution of eukaryotes in the light of phylogenomics. In: Genome Biology. Band 11, Nr. 5, S. 209, 5. Mai 2010; doi:10.1186/gb-2010-11-5-209, PMID 20441612, PMC 2898073 (freier Volltext).
- ↑ Tom Cavalier-Smith: Archaebacteria and Archezoa. In: Nature. 339. Jahrgang, Nr. 6220, Mai 1989, S. 100–101, doi:10.1038/339100a0, PMID 2497352.
- ↑ Patrick J. Keeling: A kingdom’s progress: Archezoa and the origin of eukaryotes. In: BioEssays. Band 20, Nummer 1, Januar 1998, S. 87–95, doi:10.1002/(SICI)1521-1878(199801)20:1<87::AID-BIES12>3.0.CO;2-4 (alternativer Volltextzugriff: SemanticScholar).
- ↑ William F. Martin, Miklós Müller: The hydrogen hypothesis for the first eukaryote. In: Nature. 392. Jahrgang, Nr. 6671, 1998, S. 37–41, doi:10.1038/32096, PMID 9510246.
- ↑ Hydrogen hypothesis. Auf ChemEurope.com, Lumitos AG.
- ↑
Daniel B. Mills, Richard A. Boyle, Stuart J. Daines, Erik A. Sperling, Davide Pisani, Philip C. J. Donoghue, Timothy M. Lenton: Eukaryogenesis and oxygen in Earth history. In: Nature Ecology & Evolution, 21. April 2022; doi:10.1038/s41559-022-01733-y. Dazu:
- Neue Studie legt nahe, dass eukaryotische Zellen in anoxischer Umgebung entstanden sind. Auf: Aus Der Welt (Max) vom 27. April 2022.
- Eukaryotic Cells Emerged in Anoxic Environment, New Study Suggests. Auf: sci-news vom 27. April 2022. Im Bild: Prometheoarchaeum syntrophicum MK-D1.
- ↑ Christa Schleper, Filipa L. Sousa: Meet the relatives of our cellular ancestor. In: Nature, Band 577, 15. Januar 2020, S. 478-479; doi: 10.1038/d41586-020-00039-y.
- ↑ Elizabeth Pennisi: Tentacled microbe hints at how simple cells became complex. In: Science, Band 365, Nr. 6454, 16. August 2019, S. 631; doi:10.1126/science.365.6454.631, PMID 31416944.
- ↑ Purificación López-García, David Moreira: The Syntrophy hypothesis for the origin of eukaryotes revisited. In: Nature Microbiology, Band 5, 17. April 2020, S. 655–667; doi:10.1038/s41564-020-0710-4, PMID 32341569, Epub 27. April 2020.
- ↑
Laura Eme, Daniel Tamarit, Eva F. Cáceres, Courtney W. Stairs, Valerie De Anda, Max E. Schön, Kiley W. Seitz, Nina Dombrowski, William H. Lewis, Felix Homa, Jimmy H. Saw, Jonathan Lombard, Takuro Nunoura, Wen-Jun Li, Zheng-Shuang Hua, Lin-Xing Chen, Jillian F. Banfield, Emily St. John, Anna-Louise Reysenbach, Matthew B. Stott, Andreas Schramm, Kasper U. Kjeldsen, Andreas P. Teske, Brett J. Baker, Thijs J. G. Ettema: Inference and reconstruction of the heimdallarchaeial ancestry of eukaryotes. In: Nature, Band 618, S. 992–999; doi:10.1038/s41586-023-06186-2. Dazu:
- Nadja Podbregar: Wir sind alle „Asgardianer“ – Eukaryoten entstanden aus einer Untergruppe der Asgard-Archaeen. Auf: scinexx.de vom 26. Juni 2023.
- Asgard Archaea and All Eukaryotes Share Common Ancestor, Study Says. Auf Sci.News vom 26. Juni 2023.
- ”We’re All Asgardians”: Scientists Discover New Clues About the Origin of Complex Life. Auf: SciTechDaily vom 28. Juli 2023.
- ↑ a b Patrick Forterre: Evolution - Die wahre Natur der Viren. In: Spektrum August 2017, S. 37-41 (Online-Artikel vom 19. Juli 2017)
- ↑ Tokyo University of Science: New giant virus may help scientists better understand the emergence of complex life - Large DNA virus that helps scientists understand the origins of DNA replication and the evolution of complex life In: EurekAlert!, 30. April 2019. Abgerufen am 2. Juli 2019
- ↑ Pseudomonas phage 201phi2-1, auf: Virus-Host DB, TAX:198110.
- ↑ Proteomes - Pseudomonas phage 201phi2-1, auf: UniProt.
- ↑ Vorrapon Chaikeeratisak, Katrina Nguyen, Kanika Khanna, Axel F. Brilot, Marcella L. Erb, Joanna K. C. Coker, Anastasia Vavilina, Gerald L. Newton, Robert Buschauer, K. Pogliano, E. Villa, D. A. Agard, Joe Pogliano: Assembly of a nucleus-like structure during viral replication in bacteria. In: Science. 355. Jahrgang, Nr. 6321, 13. Januar 2017, S. 194–197, doi:10.1126/science.aal2130, PMID 28082593, PMC 6028185 (freier Volltext), bibcode:2017Sci...355..194C.
- ↑
Thomas G. Laughlin, Amar Deep, Amy M. Prichard, Christian Seitz, Yajie Gu, Eray Enustun, Sergey Suslov, Kanika Khanna, Erica A. Birkholz, Emily Armbruster, J. Andrew McCammon, Rommie E. Amaro, Joe Pogliano, Kevin D. Corbett, Elizabeth Villa: Architecture and self-assembly of the jumbo bacteriophage nuclear shell. In: Nature, 3. August 2022; doi:10.1038/s41586-022-05013-4. Dazu:
- Tessa Koumoundouros: Giant Viruses Called 'Jumbo Phages' Could Help Us Fight Antibiotic Resistance. Auf: sciencealert vom 5. August 2022.
- ↑ Vorrapon Chaikeeratisak, Katrina Nguyen, M. E. Egan, Marcella L. Erb, Anastasia Vavilina, Joe Pogliano: The Phage Nucleus and Tubulin Spindle Are Conserved among Large Pseudomonas Phages. In: Cell Reports. 20. Jahrgang, Nr. 7, 15. August 2017, S. 1563–1571, doi:10.1016/j.celrep.2017.07.064, PMID 28813669, PMC 6028189 (freier Volltext).
- ↑
Tomas Alarcón-Schumacher, Susanne Erdmann: A trove of Asgard archaeal viruses. In: Nature Microbiology. Band 7, 27. Juni 2022, S. 931–932; doi:10.1038/s41564-022-01148-2. Dazu:
- Nicoletta Lanese: These Newly Discovered Viruses May Have Shaped The Rise of Complex Life on Earth. Auf: sciencealert vom 2. Juli 2022. Quelle: LiveScience.
- ↑
Ian M. Rambo, Marguerite V. Langwig, Pedro Leão, Valerie De Anda. Brett J. Baker: Genomes of six viruses that infect Asgard archaea from deep-sea sediments. In: Nature Microbiology. S. 953–961, Juli 2022; doi:10.1038/s41564-022-01150-8, Volltext (E-Book), Epub 27. Juni 2022. Dazu:
- Ermöglichten Viren komplexes Leben? Auf: wissenschaft.de vom 27. Juni 2022.
- Tiffany Winfrey: New Microbiology Research Provides Evidence Virus Plays a Role in Evolution of Asgard Archaeans. Auf: The Science Times, 27. Juni 2022. Anm.: Asgard ist keine class of bacteria, richtiger wäre etwa group of archaea.
- Incredible Virus Discovery Offers Clues About the Origins of Complex Life. Auf: SciTechDaily vom 27. Juni 2022.
- Ian M. Rambo, Valerie de Anda, Marguerite V. Langwig, Brett J. Baker: Unique viruses that infect Archaea related to eukaryotes. Preprint auf: CSH bioRxiv, 29. Juli 2021; doi:10.1101/2021.07.29.454249, ResearchGate.
- ↑ Daniel Tamarit, Eva F. Cáceres, Mart Krupovic, Reindert Nijland, Laura Eme, Nicholas P. Robinson, Thijs J. G. Ettema: A closed Candidatus Odinarchaeum chromosome exposes Asgard archaeal viruses. In: Nature Microbiology. 27. Juni 2022; doi:10.1038/s41564-022-01122-y.
- ↑ Sofia Medvedeva, Jiarui Sun, Natalya Yutin, Eugene V. Koonin, Takuro Nunoura, Christian Rinke, Mart Krupovic: Three families of Asgard archaeal viruses identified in metagenome-assembled genomes. In: Nature Microbiology. Band 7, S. 962–973962–973, 27. Juni 2022, doi:10.1038/s41564-022-01144-6, Volltext.
- ↑
Julian Vosseberg, Jolien J. E. van Hooff, Marina Marcet-Houben, Anne van Vlimmeren, Leny M. van Wijk, Toni Gabaldón, Berend Snel: Timing the origin of eukaryotic cellular complexity with ancient duplications. In: Nature Ecology & Evolution, 26. Oktober 2020, doi:10.1038/s41559-020-01320-z. Dazu:
Timeline Unveiled for One of the Most Important and Puzzling Events in the Evolution of Life, auf: SciTechDaily vom 4. November 2020, Quelle: Institute for Research in Biomedicine (IRB Barcelona) – ‚Bakterien‘ ist hier im Sinn von ‚Prokaryoten‘ zu verstehen. - ↑ Traci Watson: The trickster microbes that are shaking up the tree of life, in: Nature vom 14. Mai 2019 (englisch), Trickser-Bakterien schütteln den Stammbaum des Lebens, in: Spektrum.de vom 20. Juni 2019 (deutsch) – die Bezeichnung ‚Bakterien‘ ist nicht ganz korrekt, bei den betrachteten Mikroben handelt es sich um Archaeen oder (nach Ansicht mancher Forscher) jedenfalls um von den Bakterien verschiedene Proto-Eukaryonten.
- ↑
Julien Guglielmini, Anthony Woo, Mart Krupovic, Patrick Forterre, Morgan Gaia: Diversification of giant and large eukaryotic dsDNA viruses predated the origin of modern eukaryotes. In: PNAS, Band 116, Nr. 39, 10. September 2019, S. 19585-19592; doi:10.1073/pnas.1912006116, PMID 31506349. Siehe insbes. Fig. 2. Dazu:
- Preprint auf: bioRxiv vom 29. Oktober 2018, bioRxiv: 10.1101/455816v1 (Preprint-Volltext), doi:10.1101/455816.
- ↑ Eugene V. Koonin, Natalya Yutin: Multiple evolutionary origins of giant viruses. in: F1000 Research, 22. November 2018, doi:10.12688/f1000research.16248.1, version 1.
- ↑ Stuart A. Ralph, Bernardo J. Foth, Neil Hall, Geoffrey I. McFadden: Evolutionary Pressures on Apicoplast Transit Peptides. In: Molecular Biology and Evolution. 21. Jahrgang, Nr. 12, Dezember 2004, S. 2183–2191, doi:10.1093/molbev/msh233, PMID 15317876 (oup.com).
- ↑ Edouard Chatton: Pansporella perplexa. Réflexions sur la biologie e la phylogénie des protozoaires. In: Annales des Sciences Naturelles: Zoologie Sér. 10, Band 8, 1925, S. 5–84.
- ↑ Marie-Odile Soyer-Gobillard: Edouard Chatton (1883–1947) and the dinoflagellate protists: concepts and models. In: International Microbiology. Band 9, 2006, S. 173–177. (pdf) ( des vom 24. Dezember 2015 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- ↑ Carl Woese, G. Fox: Phylogenetic structure of the bacteria domain: the primary kingdoms. In: Proc Natl Acad Sci USA. 74. Jahrgang, Nr. 11, 1977, S. 5088–90, doi:10.1073/pnas.74.11.5088, PMID 270744, PMC 432104 (freier Volltext), bibcode:1977PNAS...74.5088W (englisch).
- ↑ Carl Woese, O. Kandler, M. Wheelis: Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. In: Proc Natl Acad Sci USA. 87. Jahrgang, Nr. 12, 1990, S. 4576–9, doi:10.1073/pnas.87.12.4576, PMID 2112744, PMC 54159 (freier Volltext), bibcode:1990PNAS...87.4576W (pnas.org [abgerufen am 11. Februar 2010]).
- ↑ a b John M. Archibald: The eocyte hypothesis and the origin of eukaryotic cells. In: PNAS. 105. Jahrgang, Nr. 51, 23. Dezember 2008, S. 20049–20050, doi:10.1073/pnas.0811118106 (pnas.org [abgerufen am 5. Oktober 2012]).
- ↑ James A. Lake, Eric Henderson, Melanie Oakes, Michael W. Clark: Eocytes: A new ribosome structure indicates a kingdom with a close relationship to eukaryotes. In: PNAS. 81. Jahrgang, Nr. 12, Juni 1984, S. 3786–3790, doi:10.1073/pnas.81.12.3786, PMID 6587394, PMC 345305 (freier Volltext) – (pnas.org [abgerufen am 5. Oktober 2012]).
- ↑ Steve Kelly, B. Wickstead, K. Gull: Archaeal phylogenomics provides evidence in support of a methanogenic origin of the Archaea and a thaumarchaeal origin for the eukaryotes. In: Proceedings of the Royal Society B. 278. Jahrgang, 2011, S. 1009–1018, doi:10.1098/rspb.2010.1427, PMID 20880885, PMC 3049024 (freier Volltext) – (englisch, stevekelly.eu ( des vom 3. März 2016 im Internet Archive) [abgerufen am 18. Dezember 2022]).
- ↑ Anthony M. Poole, Nadja Neumann: Reconciling an archaeal origin of eukaryotes with engulfment: a biologically plausible update of the Eocyte hypothesis. In: Research in Microbiology. 162. Jahrgang, 2011, S. 71–76, doi:10.1016/j.resmic.2010.10.002 (nordita.org [PDF; abgerufen am 5. Oktober 2012]).
- ↑ Lionel Guy, Thijs J. G. Ettema: The archaeal 'TACK' superphylum and the origin of eukaryotes. In: Trends in Microbiology. 19. Jahrgang, Nr. 12, Dezember 2011, S. 580–587, doi:10.1016/j.tim.2011.09.002, PMID 22018741 (sciencedirect.com [abgerufen am 5. Oktober 2012]).
- ↑ Cymon J. Cox, Peter G. Foster, Robert P. Hirt, Simon R. Harris, T. Martin Embley: The archaebacterial origin of eukaryotes. In: PNAS. 105. Jahrgang, 23. Dezember 2008, S. 20356–20361, doi:10.1073/pnas.0810647105, PMID 19073919, PMC 2629343 (freier Volltext) – (pnas.org [abgerufen am 5. Oktober 2012]).
- ↑ Anja Spang, Jimmy H. Saw, Steffen L. Jørgensen, Katarzyna Zaremba-Niedzwiedzka, Joran Martijn, Anders E. Lind, Roel van Eijk, Christa Schleper, Lionel Guy, Thijs J. G. Ettema: Complex archaea that bridge the gap between prokaryotes and eukaryotes. In: Nature. 521. Jahrgang, 2015, ISSN 0028-0836, S. 173–179, doi:10.1038/nature14447, PMID 25945739, PMC 4444528 (freier Volltext).
- ↑ Laura Eme, Anja Spang, Jonathan Lombard, Courtney W. Stairs, Thijs J. G. Ettema: Archaea and the origin of eukaryotes. In: Nature Reviews Microbiology. 15. Jahrgang, Nr. 12, 10. November 2017, ISSN 1740-1534, S. 711–723, doi:10.1038/nrmicro.2017.133 (englisch, nature.com).
- ↑ Katarzyna Zaremba-Niedzwiedzka, Eva F. Cáceres, Jimmy H. Saw, Disa Bäckström, Lina Juzokaite, Emmelien Vancaester, Kiley W. Seitz, Karthik Anantharaman, Piotr Starnawski, Kasper U. Kjeldsen, Matthew B. Stott, Takuro Nunoura, Jillian F. Banfield, Andreas Schramm, Brett J. Baker, Anja Spang, Thijs J. G. Ettema: Asgard archaea illuminate the origin of eukaryotic cellular complexity. In: Nature, Band 541, S. 353–358, 19. Januar 2017; doi:10.1038/nature21031.
- ↑ NCBI Taxonomy Browser: Archaea; Details: "Archaea" Woese et al. 1990 (superkingdom); graphisch: Archaea, auf: Lifemap NCBI Version.
- ↑ Bram Henneman: Histone-DNA assemblies in archaea: shaping the genome on the edge of life. Leiden University Repository, 4. Dezember 2019; hdl:1887/81191. Hier: Kapitel 1: Introduction (englisch). Siehe Fig. 1.1.
- ↑ Thomas Cavalier-Smith: The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. In: Int. J. Syst. Evol. Microbiol. 52. Jahrgang, Pt 2, März 2002, S. 297–354, doi:10.1099/00207713-52-2-297, PMID 11931142 (microbiologyresearch.org).
- ↑ Regina Saum, Katharina Schlegel, Benjamin Meyer, Volker Müller: The F1FO ATP synthase genes in Methanosarcina acetivorans are dispensable for growth and ATP synthesis. In: FEMS Microbiology Letters, Band 300, Nr. 2, November 2009, S. 230–236; doi:10.1111/j.1574-6968.2009.01785.x.
- ↑ Elena Hilario, Johann Peter Gogarten: Horizontal transfer of ATPase genes - the tree of life becomes a net of life. In: BioSystems. 31. Jahrgang, Nr. 2–3, 1993, S. 111–119, doi:10.1016/0303-2647(93)90038-E. PMID 8155843 ScienceDirect Dieser Artikel bei University of Connecticut (PDF).
- ↑ Armen Y. Mulkidjanian, Michael Y. Galperin, Kira S. Makarova, Yuri I. Wolf, Eugene V. Koonin: Evolutionary primacy of sodium bioenergetics. In: Biology Direct. 3. Jahrgang, Nr. 13, 2008, doi:10.1186/1745-6150-3-13 (englisch).
- ↑ Armen Y. Mulkidjanian, Kira S. Makarova, Michael Y. Galperin, Eugene V. Koonin: Inventing the dynamo machine: the evolution of the F-type and V-type ATPases. In: Nature Reviews Microbiology. 5. Jahrgang, Nr. 11, 2007, S. 892–899, doi:10.1038/nrmicro1767 (nature.com [PDF]). PDF (via Universität Osnabrück), Memento im Webarchiv vom 31. Oktober 2008.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Weblinks
Bearbeiten- Eukaryonten Zellaufbau
- Eukaryoten (Tree of Life Web Project) (englisch)
- Botanik online, Universität Hamburg
- Wilfried Probst: Frühe Evolution und Symbiose, Europa-Universität Flensburg, Institut für Biologie und Sachunterricht und ihre Didaktik: §LECA und Mitochondrien, abgerufen am 19. April 2019
- Masaharu Takemura: Medusavirus Ancestor in a Proto-Eukaryotic Cell: Updating the Hypothesis for the Viral Origin of the Nucleus. In: Front. Microbiol., Band 11, 571831, 3. September 2020. doi:10.3389/fmicb.2020.571831. Dazu:
Tessa Koumoundouros: Giant Viruses Could Explain The Mysterious Evolution of a Key Part of Our Cells, auf: sciencealert vom 10. September 2020
In Ancient Giant Viruses Lies the Truth: Medusavirus Key to Deciphering Evolutionary Mystery, auf: SciTechDaily vom 11. September 2020 - Ancestors of Legionella Bacteria – Which Causes Legionnaires’ Disease – Infected Cells Two Billion Years Ago. Auf SciTechDaily vom 15. Februar 2022. Quelle: Universität Uppsala.
- Lenka Horváthová et al.: Analysis of diverse eukaryotes suggests the existence of an ancestral mitochondrial apparatus derived from the bacterial type II secretion system. In: Nature Communications, Band 12, Nr. 2947, 19. Mai 2021; doi:10.1038/s41467-021-2304. Siehe Sec-System und Bakterielle Proteinsekretion §Typ II.